лни юпухоекюц цскюц - MY GULAG ARCHIPELAGO (38)
Comparative morphology of the palate dentition in Proganochelys quenstedti Baur 1887 from the Upper Triassic of Germany and chelonian ancestry
Elena G. Kordikova, Tuebingen
PDF in process
Introduction
The Chelonia form a well-defined group of tetrapods which is mainly characterized by the development of a shell and the derivation of a stegocephalian skull. A saltatory formation of a chelonian shell in the geological record, which led to the migration of the limb girdles to positions inside the rib cage, has caused a big interest to the questions connected with the formation of the shell and origin of the group. The existence of a temporal and morphological hiatus between most turtles, on the one hand, which lost teeth but already had a shell, and their presumable ancestors among Paleozoic and Mesozoic tetrapods, on the other hand, having teeth but didn▓t developed a shell of chelonian type made the chelonian origin one of the most fascinating problems in herpetology and paleoherpetology.
Hypotheses of the turtle origin
At present there are a dozen of different hypotheses connected with origin and evolution of the group. In the past turtles were considered to be sister taxa to: ichthyopterygians (Baur 1887), diadectomorphs (Huene 1912, 1948), etc. A first discussion of the chelonian relationships in modern terms was made by Gregory (1946) who recognized the close similarity between turtles, placodonts and pareiasaurs. Ivakhnenko (1987) and Laurin and Reisz (1995) ally the ancient turtle Proganochelys with procolophonoids rather than with pareiasaurs. Ivakhnenko also suggests that Proganochelys is not a chelonian but a very specialized turtle-like tetrapod. However, according to the traditional point of view and last data, Proganochelys has to be considered as a terrestrial, well-armoured turtle within Chelonia.
Later, Lee (1993, 1995, 1996, 1997a,b) developed and reinforced arguments for the pareiasauroid origin of Chelonia.
However, the chelonian relationships with the different outgroups which had been suggested earlier as ancestors, have not been tested. Taken into account a general morphology of Proganochelys Gaffney (1990) suggests that the turtles may have evolved from the early Captorhinida. Gauthier et al. (1988) also consider Chelonia as a sister taxon of captorhinids and classify them together with diapsids and their presumed fossil relatives (Paleothyris) in the Reptilia. Rieppel and DeBraga (1996), DeBraga and Rieppel (1997) and Rieppel (1999) consider turtles as a sister-group of the Sauropterygia, both nested at the base of the lepidosauromorph lineage. There is also a lot of molecular works linking turtles with lepido- and archosauromorphs (e.g., Zardoya & Meyer 1998, Hedges & Poling 1999, etc.).
Temporal and jugal emarginations
Tetrapods are usually classified on the basis of the pattern of openings in the dermal skull roof behind the orbits. In primitive amniotes, this area is completely covered with bone. This pattern is termed anapsid and forms the basis for the subclass Anapsida (Fig. 1).
|
|
Fig. 1. Skulls of early amniotes showing the patterns of temporal fenestration.
|

The turtles are usually considered among the anapsids since they retain a continuous bony covering behind the orbits and have no temporal arcades (Osborn 1903, Williston 1904, Romer 1956, Carroll 1988). However, some authors have proposed that the anapsid turtle skull developed secondarily from skull patterns with one or more pairs if temporal openings (Broom 1924, Goodrich 1930, DeBeer 1937, Rieppel & DeBraga 1996). The first group to diverge from the ancestral stock were the synapsids. This group, including the mammal-like reptiles, has a single pair of openings located low in the cheek and bordered by the jugal, squamosal, and postorbital. Late in the Pennsylvanian, a second major group di╜verged from the basal anapsid stock: the Diapsida. Diapsids are characterized by the presence of two pairs of temporal openings - one pair located ventral to the postorbital and a second pair dorsal to the postorbital and squamosal and lateral to the pa╜rietal. It is considered this stock gave rise to all living reptiles subdivided into the Lepidosauromorphs and Archosauromorphs. Two major groups of Mesozoic marine reptiles, the plesiosaurs and ichthyosaurs, have an upper temporal opening like that of the diapsids but lack a clearly defined lateral opening. This pattern has been termed parapsid or euryapsid and was thought to have arisen directly from an anapsid configuration or can be derived from that of early diapsids by elimination of the lower temporal bar and thickening of the postor╜bital and squamosal (e.g., Carroll 1988).
The earliest fossil turtle, Proganochelys from the Upper Triassic of Germany, demonstrate a closed temporal region (Fig. 2).
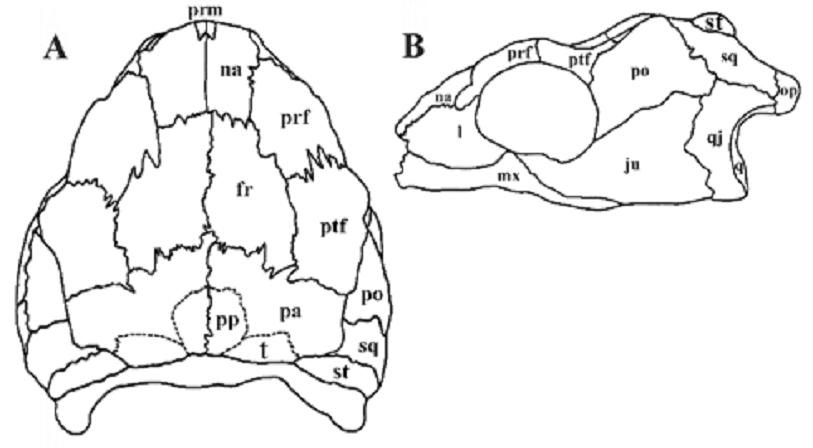 |
Fig. 2. Skull of Proganochelys quenstedti Baur 1887. After Kordikova (2002). Dorsal (A) and ventral (B) views. Abbreviations: ju - jugal, l - lacrimal, mx - maxilla, na - nasal, pa √ parietal, po - postorbital, prf - prefrontal, prm - premaxilla, ptf - postfrontal, pt - pterygoideum, q - quadrate, qj - quadratojugal, sq - squamosal, st - supratemporal, t - tabular. |
Absence of fenestration in basal chelonians can be most reasonably interpreted in only two ways:
1. Proganochelys evolved from synapsid- or diapsidlike reptiles by secondary closure of fenestra which appeared in different lineages of descendants; or
2. Proganochelys evolved from parareptiles not having any fenestration and the ancient turtles just retained this condition.
The first hypothesis requires a character reversal in the line leading to Proganochelys. It also implies that an ancestral morphogenetic program has to be realized in different groups of Chelonia. However, the manner of emargination of the skull roof in turtles is completely different from that in synapsid- or diapsidlike reptiles (Kordikova 2002). Analysis of possibilities of the temporal fenestration in different groups of tetrapods made by FrazZetta (1968) also confirms this point of view. The skull morphology is now the focus of a renewed effort to treat the evolution of the skull patterns in Chelonia. Although this work is still in preparation (Kordikova personal investigations) the scheme of the skull evolution would be useful to demonstrate that the skull of turtles recapitulates neither the diapsid pattern of Archosauromorpha (as Rieppel & DeBraga 1996 suggest) nor synapsid one but derives from a stegocephalian type. According to preliminary investigations of skulls in fossil and Recent turtles I suppose that during evolution of the stegal skull pattern in Chelonia took place either as progressive emargination (jugal or temporal) of the skull roof or retention of the anapsid pattern, but not a skull fenestration. In other words, ⌠true■ windows like in synapsid- or diapsidlike reptiles are not formed in the chelonian skull. The jugal emargination proceeded from ventrolaterally to dorsoposteriorly and led to the complete underdevelopment of quadratojugal and the partial underdevelopment of the temporal bones including the parietal in ontogeny of several groups of Chelonia (e.g., chelids, Terrapene, Indotestudo, etc.) while the jugal emargination in diapsids proceeded dorsally from the jugal and quadratojugal, posteroventrally from the postorbital and anteriorly or anterodorsally from the squamosal with the formation of the lower opening (Fig. 1). The temporal region is well-developed in some fossil turtles like meiolaniids which have squamosal horns elongated posteriorly (Gaffney 1983). Temporal emargination in most turtles starts posteriorly from a reduction of the parietal, supraoccipital and squamosal and proceeds anteriorly, leading to the complete underdevelopment of the postorbital in ontogeny of Kinixys in contrast to the upper opening taking place in diapsids and synapsids. In these groups the upper opening usually remains due to the delay of ossification of the lateral part of the parietal, anterodorsal part of the squamosal and dorsoposterior part of the postorbital.
The second hypothesis is more probable. It explains the progressive emargination of the skull roof from more complete and complicated roofing with a larger set of dermal bones to the lesser one (so, the jugal and temporal emargination already took place in the Middle-Late Jurassic turtles). The anapsid configuration in turtles appears to have evolved separately from the diapsid and synapsid condition. There is no evidence for the prior existence of temporal openings in ancient turtles, which precludes their close relationship to early synapsids or diapsids. The absence of any clear-cut association between Proganochelys and archosauromorphs, e.g. the temporal fenestra, does not allow to consider these groups together.
In other words, the complete dermal ossification of the skull is a plesiomorphic character for Chelonia while the underdevelopment of different skull regions is a derived condition. The stegal type of skull, the specific morphology of the palate (presence of palate toothbearing ridges, the ectopterygoid, etc.), ventro-lateral migration of the paroccipital process, separation of the exoccipital and opisthotic, etc. are features that occur in ancient chelonians are shared rather with the procolophonoid-pareiasauroid stock than the archosauromorph lineage.
To sum, the morphological strategies in the evolution of temporal openings seem to be quite different in the parareptilian and eureptilian stocks. Taking into account the morphology of the temporal region in lower tetrapods, ancient turtles of the proganochelyid group appear to be related to the parareptilian stock than to the eureptilian one. It is obvious that modern analyses of DNA data as well as comparative anatomical information taking into account only living taxa can not elaborate turtle phylogeny.
Moreover, the assignment of the turtles to the Anapsida is somewhat arbitrary, since their ancestors probably diverged from the primitive procolophonoid - pareiasaur parareptilian stock earlier than either the synapsids or the diapsids. In addition, their overall anatomy and way of life is so different from that of the early anapsids that it is more logical to place them in a distinct subclass.
Palate morphology of Proganochelys quenstedti Baur 1887 from the Upper Triassic of Germany and the chelonian ancestry
The morphology of the palate area is quite important for respiratory and digestive adaptations in tetrapods. As known palatal dentition is absent in most turtles - Casichelydia sensu Gaffney 1975, but it is present in ancient, well-armoured chelonians (Proganochelys Baur 1887 and Kayentachelys Gaffney et al. 1987). This makes them similar morphologically to early tetrapods. Up to now a substantial structural and temporal hiatus exists between any presumed ancestral group and ancient turtles. The presence of teeth in the palate region in Proganochelys appears to be the only common link between Casichelydia which have a shell but no the palate teeth and the turtle ancestors which presumably had the palate teeth but no shell of chelonian type. Thus, the study of the palate region of the Late Triassic turtle Proganochelys and the different ancestral groups can throw light on the origin of Chelonia, forward our understanding of chelonian relationships with different outgroups and solving the question on the position of turtles within the lower tetrapod phylogeny.
Studies of the palate area and skull morphology in Proganochelys quenstedti Baur 1887 from the Upper Triassic of Germany reveal the following features (in addition to Jackel 1918, Gaffney 1990) (Fig. 3):
-
the presence of ridge thecodont palate dentition,
- premaxillary tooth vestiges,
- columnar ectopterygoid,
- additional bones in the parietal area - postparietals and tabulars,
-
different skull bone arrangement (enlarged postorbital, etc. ).
|
|
Fig. 3. Proganochelys quenstedti Baur 1887. After kordikova (2002). Palate region of the skull, general view: A -increased premaxillary area, premaxillary tooth vestiges are shown by asterisks; B,C - increased palatino-pterygoid area. Abbreviations: bptj - basipterygoid joint, bsp - basisphenoid, ch - choanae, ect - ectopterygoid, fpp - foramen palatinum posterius, ipv - interpterygoid vacuity, pal - palatine, pltr - palatine tooth ridges, pptp - processus pterygoideum posterius, pttr - pterygoid tooth ridges, v - vomer, vtr - vomerine tooth ridges.
|

Palate morphology in early tetrapods
Comparative analysis of skull morphology and, especially, of palate area in different lineages of Paleozoic tetrapods (Fig. 4) confirms the parareptilian status of ancient turtles (see also Table).
|
|
Fig. 4. Palate morphology in early tetrapods. Skull, ventral view: A. Paleothyris, B. Diadectus, C. Petrolacosaurus, D. Captorhinus, E. Nyctiphruretus, F. Bashkyroleter, G. Scutosaurus, H. Procolophon, I. Euparkeria, J. Stagonolepis, K. Placodus, L. Proganochelys, M. Chasmatosaurus, N. Sphenodon.
|
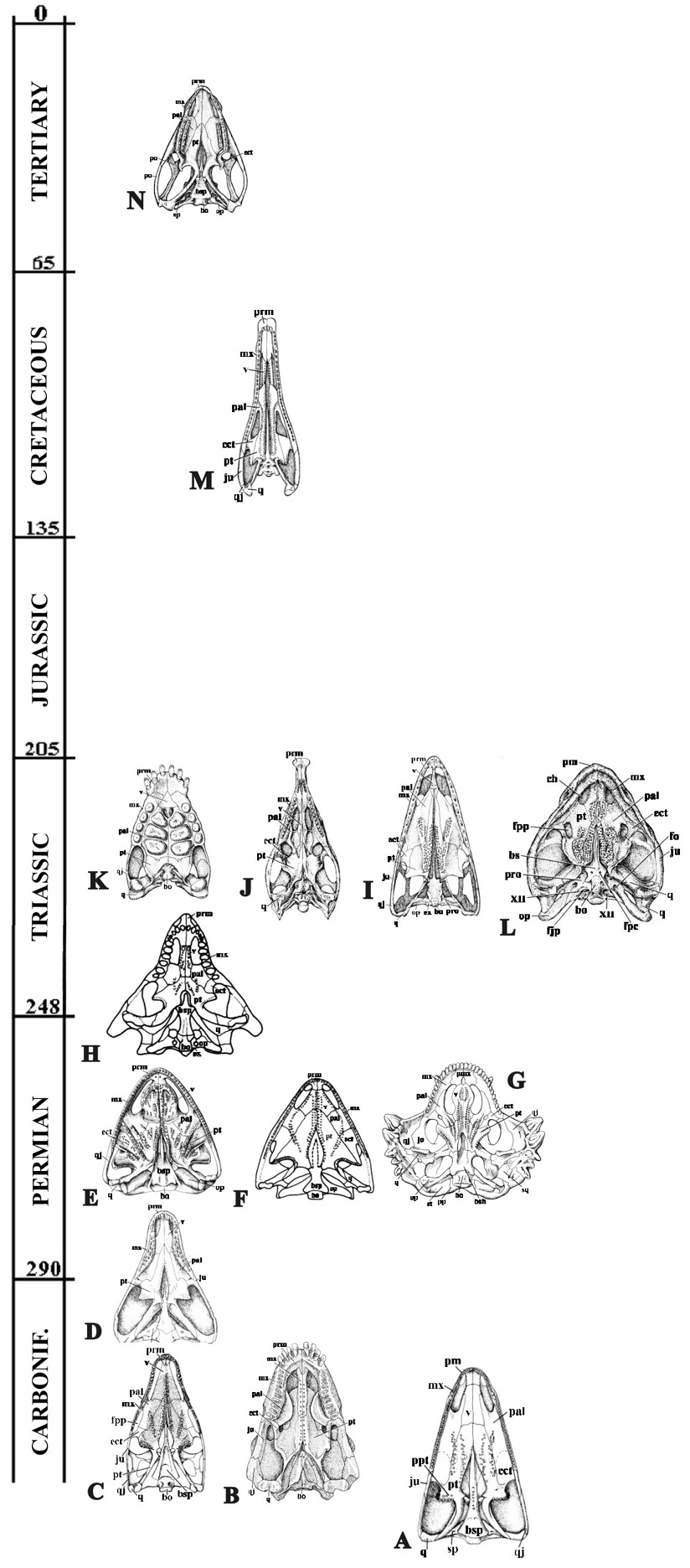
Table. The distribution of characters based mainly on the morphology of the palate area of lower tetrapods
Character definitions for Table. For each character the most primitive state is number 0. 1. palate dentition present (0) or absent (1); 2. maxillar and dentary dentition present (0) or absent (1); 3. palate teeth acrodont (0), hyperacrodont (1), prothecodont (2), pseudoacrodont (3), thecodont (4), thecodont arranged on ridges (5), or teeth absent (6); 4. palate teeth arranged into tooth fields (0), tooth-covered stripes (1), tooth ridges (2), or teeth absent (3); 5. dentition on the vomer, pterygoid, palatine (0), on the pterygoid and palatine (1), on the vomer and pterygoid (2), on the palatine (3), on the pterygoid (4) or teeth absent (5); 6. teeth on basisphenoid present (0) or absent (1); 7. teeth on transverse pterygoid flange present (0) or absent (1); 8. palate dentition isodont (0), heterodont (1) or teeth absent (2); 9. palate teeth conical (0), tricuspids (1), plate-liked (2) or teeth absent; 10. ectopterygoid present (0) or absent (1); 11. interpterygoid vacuity present (0) or absent (1); 12. foramen palatinum posterius present (0) or absent (1); 13. basipterygoid articulation present during whole life (0), present in young specimens and sutured closed in adults (1), or absent during whole life (2). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
However, Proganochelys cannot be derived directly from primitive procolophonoids that have a generalized palate pattern, on the one hand, and reduced skull structure, on the other hand.
The Proganochelys pattern also cannot be derived directly from the pareiasaurid one, which is characterized by a quite specialized skull morphology (extreme expansion of the cheek area, reduced number of palatine and pterygoid tooth ridges, sutured basipterygoid contact, presence of transverse pterygoid tooth ridge, retention of palate vacuities, etc.).
Paedomorphosis and ancestry of turtles
|
It is known that heterochrony and, in particular, paedomorphosis, quite often occurs among different lineages of Chelonia (Kordikova 2000). A reanalysis of skull morphology and palate area in Proganochelys reveals significance of paedomorphosis for the chelonian ancestry.
Many cranial features in Proganochelys quenstedti resembles closely that of young specimens of some of described pareiasaurs as Deltavjatia that is known from numerous ontogenetic series, than of adults.
During development of Proganochelys and young specimens of pareiasauroids the following trends take place:
All above-mentioned peculiarities provide evidence for a paedomorphic origin of the ancient turtle skull pattern from that of a basal pareiasauroid lineage - from tetrapods with a more complete set of skull elements and marginal dentition. Similarities in the morphology of the palate area and palate dentition in young pareiasaurids and Proganochelys assume their similar food adaptations. |
Remarks to the evolution of tooth palate attachment in lower tetrapods
The observation of ridge thecodont palate dentition in Proganochelys quenstedti has stimulated a research of patterns of palate of lower tetrapods the palate tooth attachment developed from acrodont to thecodont through a tooth attachment and their evolution in different groups of Paleozoic and Mesozoic tetrapods.
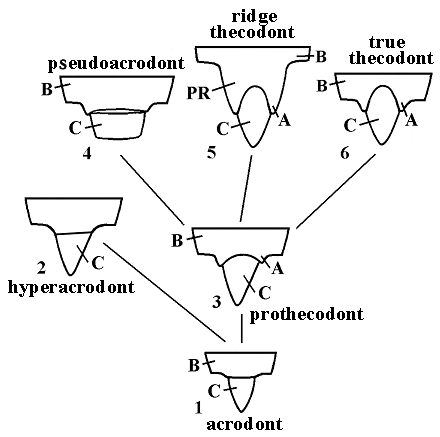
Different types of palate tooth attachments (acrodont, hyperacrodont, prothecodont, pseudoacrodont, ridge thecodont and true thecodont) are regarded (Fig. 5).
|
Fig. 5. Evolution of palate tooth attachment in tetrapods. The main types of palate dentition: 1. acrodont (Captorhinus, Diadectus), 2. hyperacrodont (Sphenodon), 3. prothecodont (early diapsids, early thecodonts, early procolophonoids), 4. pseudoacrodont (Placodus), 5. ridge thecodont (late procolophonoids, pareiasauroids, Proganochelys), 6. true thecodont (thecodont diapsids). Abbreviations: A - alveolar, B - bone tissue, C - tooth crown, PR - palatal ridge. |
The acrodont tooth attachment presented in primitive Late Paleozoic tetrapods like diadectomorphs, captorhinomorphs, early procolophonoids is considered as the most primitive state.
The palate teeth in late procolophonoids, pareiasauroids and ancient turtles are characterized by primitive ridge thecodont implantation in which the palate teeth are set in sockets and distinguished from the surrounded bone. But in contrast with the true thecodont mode, palate teeth in late parareptiles are attached to the high and well-developed tooth ridges. The tooth system in Proganochelys appears to derive from the prothecodont type as in early procolophonoids and diapsidomorphs but evolved towards ridge thecodonty together with that of late procolophonoids and pareiasauroids (Fig. 5.3,5).
The prothecodont tooth implantation also produced hyperacrodont mode of tooth attachment (Howes & Swinnerton 1901) that is characteristic for Sphenodon and pseudoacrodont one - for Placodus. Fused tooth margins and the tooth base that is not attached to the bone completely characterize pseudoacrodont type of progressive acrodont fashion in contrast with proper acrodont and prothecodont patterns. The pleurodont type of palate dentition appears to be absent in lower tetrapods because of the general construction of a flattened palate.
The form of the palate teeth in lower tetrapods can change from conical to tricuspid in pareiasaurids and to plate-like in placodontids. It is assumed that the morphology of the palate dentition in adult tetrapods reflects the early stages of tooth morphogenesis of the marginal dentition.
more information:
|
Classification of lower tetrapods Class Reptilia Linneus 1758 Subclass Parareptilia Olson 1947
Order Procolophonomorpha
Suborder
Procolophonia
Superfamily
Procolophonoidea Family Nyctiphruretidae Efremov 1938 Family Procolophonidae Seeley 1888
Suborder
Pareiasauroidea Family Lanthanosuchidae Efremov 1946 Family Nycteroleteridae Romer 1956 Family Pareiasauridae Seeley 1888 Family Bradysauridae Huene 1948 Order Chelonia brongniart 1800 Suborder Proganochelydia ROMER 1966 Suborder Casichelydia GAFFNEY 1975
Subclass Eureptilia
Order Diadectida
Suborder
Diadectomorpha
Family Diadectidae
Order Captorhinida
Suborder
Captorhinomorpha
Family Protorothyrididae
Family
Captorhinidae
Case 1911
Order Araeoscelida
Family Petrolacosauridae
Order Choristodera
Family Champsosauridae
Infraclass
Lepidosauromorpha
Superorder Lepidosauria
Order Sphenodontida
Family Sphenodontidae
Superorder Sauropterygia
Order
Placodontia
Cope
1871 Suborder Placodontoidea Cope 1871 Family Placodontidae Cope 1871 Family Paraplacodontidae Peyer & Kuhn-Schnyder 1955
Suborder
Cyamodontoidea
Nopcsa
1923 Superfamily Cyamodontida Romer 1956 Family Henodontidae F.v. Huene 1948
Family
Cyamodontidae
Nopcsa
1923
Infraclass
Archosauromorpha
Superorder Archosauria
Order Thecodontia
Suborder
Proterosuchia
Family Proterosuchidae
Suborder
Ornithosuchia
Family Euparkeriidae
Suborder
Aetosauria
Family Stagonolepididae
Order Chelonia brongniart 1800 (Latreille 1800) Suborder Proganochelydia Romer 1966 Family Proganochelyidae Baur 1888 Suborder Casichelydia GAFFNEY 1975 Infraorder Cryptodira Cope 1868 Family Pleurosternidae Cope 1868 Family Kallokibotionidae Nopcsa 1923 Family Solemydidae Lapparent de Broin & Murelaga 1996 Family Thalassemydidae Zittel 1889 Family Eurysternidae Dollo 1886 Family Plesiochelyidae Baur 1888 Superfamily Chelonioidea Oppel 1811 Family Osteopygidae Zangerl 1953 Family Protostegidae Cope 1872 Superfamily Dermochelyoidea Fitzinger 1843 Family Dermochelyidae FITZINGER 1843 Superfamily Trionychoidea Fitzinger 1826 Family Trionychidae Fitzinger 1826 Family Cyclanorbidae LYDEKKER 1889 Family Peltochelyidae Seeley 1880 Superfamily Carettochelyoidea Boulenger 1887 Family Carettochelydidae Boulenger 1887 Family Kinosternidae Agassiz 1857 Family Staurotypidae GRAY 1869 Superfamily Chelydroidea gray 1831 Family Platysternidae gray 1869 Family Dermatemydidae gray 1870 Superfamily Testudinoidea batsch 1788 Family Testudinidae batsch 1788 Superfamily Emydoidea Rafinesque 1815 Family Emydidae Rafinesque 1815 Infraorder Pleurodira COPE 1864 Family Proterochersidae Nopcsa 1928 Family Platychelyidae BrДm 1965 Family Dortokidae Lapparent de Broin & Murelaga 1996 Superfamily Pelomedusoides Cope 1868 Family Pelomedusidae Cope 1868 Superfamily Podocnemidoidea Cope 1868 Family Bothremydidae Baur 1891 Family Podocnemididae Cope 1868 Superfamily Chelidoidea Gray 1825
|