╠╬╔ └ąš╚Ž┼╦└├ ├ė╦└├ - MY GULAG ARCHIPELAGO (38)
Heterochrony in the evolution of the shell of Testudinata. Part 1. Nomenclature. Cheloniidae, Dermochelyidae, Trionychoidea (Trionychidae, Cyclanorbidae) and Carettochelyidae
Elena G. Kordikova, Tuebingen
PDF in process
1. Bony shell
1.1. Carapace
|
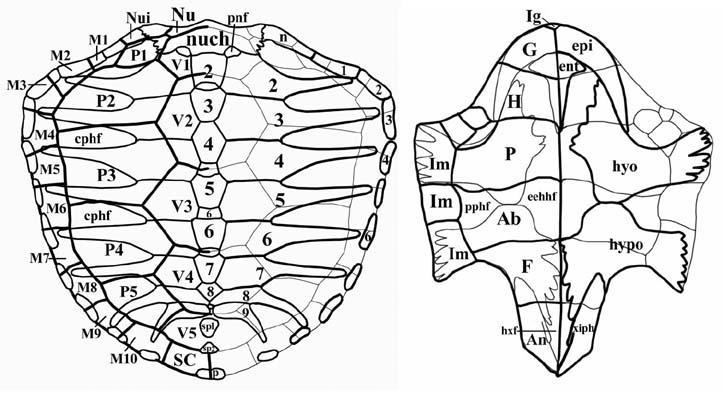
Caretta caretta (L. 1758), Cheloniidae
Ab - an abdominal scute, An - an anal
scute, Ax ¢ an axillary scute, cphf - a
costoperipheral fontanelle, EEHH ¢
epientohyoplastral hinge, EHH -
epihyoplastral hinge, ent -
entoplastron; epi - epiplastron; F - a
femoral scute, G - a gular scute, H - a
humeral scute, hhf ¢ a hyohypoplastral
fontanelle, hhxf - a
hyohypoxiphiplastral fontanelle, hxf -
hypoxiphiplastral fontanelle, hyo -
hyoplastron; hyp ¢ hypoplastron, HXH -
hypoxiphiplastral hinge, Ia ¢ an
interanal scute, Ig - an intergular
scute, Im - an inframarginal scute, M ¢
a marginal scute, msp - mesoplastron, n
- a peripheral contacting with a nuchal,
Nu - a nuchal scute, nuch - a nuchal,
Nui - a nuchaloid scute, p - a pygal;
pepi - preepiplastron, prn - a
prenuchal, SC ¢ supracaudal scute, sp -
a suprapygal, ppf ¢ plastroperipheral
fontanelle, xiph - xiphiplastron, xf - a
xiphiplastral fontanelle. |
Ę Prenuchal ¢ a small oval bone at the front of the carapace in some trionychoids - cyclanorbids (Lissemys spp., Cyclanorbis senegalensis) and trionychids (Dogania subplana, plastomenusines, etc.). It occurs sporadically in old specimens of the trionychine Platypeltis ferox.
Ę Nuchal - a large, wide bone at the front of the carapace. This name is nearly universal although Carr (1952) called it the ōproneuralö.
Ę
Neurals ¢
the series of bones behind the nuchal outgrowths of
the neural arches of the thoracic vertebrae (see
discussion in
Kordikova (2000). The term is
universally accepted in spite of some authors also
using ōvertebral platesö. Besides, many researchers
use the term äpreneuralō for the most anterior
neural if it is highly differentiated in form or
ōextraö. However, this term just demonstrates the
specific status and position of the plate. It is not
so informative because it does not facilitate
homologization of thoracic vertebrae of turtles with
those of other tetrapods. A number of authors (Hasan
1941,
Carpenter (1981,
Meylan 1984,
Kordikova
1993b, 2000) call the most anterior neural
(äpreneuralō) äthe first neuralō. Two neurals
between a first pair of costals occur as a norm in
some trionychoids (Lissemys spp.,
cyclanorbines, aspideretines, Late Cretaceous
paraplastomenusines, Late Cretaceous and Eocene
ulutrionychines), baenids, etc. This bone in
above-mentioned turtles is not sutured to the neural
arch of the thoracic vertebra I but appears to be
formed as an expansion of perichondral ossifications
of the neural arch of the thoracic vertebra I (Kordikova
1993b, 2000; compare with
Cherepanov
1995). The neural I is reduced in most chelonians
but it is not fused with next to it neural as
Webb
(1962),
Gaffney (1979) and
Meylan
(1987) suggested. I also prefer to call äfirst
neuralō the most anterior neural, taking into
account its formation as a modified element of the
neural arch of the thoracic vertebra I (Kordikova
1991, 1993b, 2000). The following neurals after the
neural I have corresponding numeration: the neural
of the thoracic vertebra II is the neural II, the
neural of the vertebra III is the neural III, etc.
Neurals are often
reduced or missing in some chelids,
cyclanorbines, etc.
Ę
Costals ¢
the series of bones on each side of the neurals.
Many authorities use this term, although some
authors use "pleurals" (Meylan
1987,
Gaffney 1990,
Pritchard & Trebbau 1984,
etc.).
Pritchard
&
Trebbau (1984) take into consideration the
position of bones, which are more superficial
structures, external to the lungs or pleura
and argue that it is preferable to use an
exclusively Greek nomenclature for the carapace
bones and Latin nomenclature for the scutes.
However, I prefer to use äcostalsō because this term
comes from the ribs or costae and points to the
origin of the plates, which form from outgrowths of
corresponding ribs. Moreover, costals are modified
ribs of the corresponding vertebrae and,
consequently, the rib of the thoracic vertebra I
forms its own costal I, the rib of the vertebra II
forms the costal II, etc.
(Kordikova
2000). Usage of
either Greek or Latin term is inessential if it does
not reflect homology of bony elements and confuses a
number of researchers. Some authors (Siebenrock
1902,
Meylan 1984, 1987) use a term äcallositiesō
for the carapace and plastral ossifications that
refers to the dermal origin of the costals.
Ę
Peripherals
- bones forming the edge of the carapace, a
nearly universal term. However, they are sometimes
termed "marginals." Peripherals are formed by
respective myosepta, which also take part in the
formation of corresponding ribs and vertebrae (Cherepanov
1989). Thus, the numbering of peripherals
corresponds to that of the myosepta and,
consequently, the ribs and vertebrae: the myoseptum
number I is that from which the thoracic rib (I) and
peripheral (I) originates, the myoseptum II is that
from which the thoracic rib (II) and peripheral (II)
originates, etc. The nuchal and the nuchal
peripherals also originate in the same myoseptum (Kordikova
2000).
Ę
Pygal
is the most posterior bone of the carapace
forming together with peripherals the edge of the
carapace; a term universally used.
Ę
Suprapygals
- the bones between the neurals and the pygal,
of which suprapygals may be one or more. In
comparison with neurals suprapygals are formed in
the dermis independently but not as outgrowths of
the neural arches of the thoracic vertebrae. They
have a sutured contact with vertebrae secondarily (Cherepanov
1989).
Boulenger (1889) called these bone the
ōpygalsö, but otherwise ōsuprapygalö is virtually
unanimous.
1.2. Plastron
The paired bones of the plastron are, from anterior to posterior, the epiplastra,
hyoplastra,
hypoplastra, and xiphiplastra. A single
bone encircled by the epiplastra and hyoplastra is
the entoplastron (Smith
& Smith 1980, preferred the spelling
"endoplastron"). The additional bones between the
hyoplastra and hypoplastra in proganochelyids,
platypeltids, baenids, desmostylids, pelomedusids,
etc. are the mesoplastra. Sometimes they are
called interplastra (Sukhanov
1964). An additional pair of
epiplastral bones ¢ preepiplastra can form in
the cyclanorbid Cyclanorbis senegalensis.
There are many authors
(Siebenrock
1902, Romer 1956,
Zangerl
1969,
Meylan 1984, 1987, etc.) who distinguish two
kinds of plastral bones in the chelonian shell and
use a term äcallositiesō for plastral ossifications.
However, such viewpoint is not confirmed by
morphogenesis - there are no independent superficial
ossicles over the plastral bony primordial (see wide
discussion on this question in
Cherepanov
1997 as well as in
Kordikova
2000). Here, I prefer to use plastral bones or
plastral ossifications for the epi-, hyo-, hypo-,
xiphiplastra and entoplastron.
2. Horny shell
2.1 Carapace
I use terms ōscaleö
and ōscuteö for shell integumentary structures:
ōscalesö for tiny horny formations distributed on
the whole body, including shell, and ōscutesö for
the wide horny plates forming the surface of the
shell. Scales are present in all turtles (they are
ossified in dermochelyids) while scutes are present
in most chelonians except trionychoids and
dermochelyids (scutes are usually reduced in
chelonioids and carettochelyids).
Carr
(1952) proposed that scutes be termed "laminae"
instead, but
Zangerl (1969) observed correctly that this
term is ambiguous, having an established usage in
osteology, and should not be used. The old term
ōcorneoscutesö is accurate but laborious and is
little used today (Pritchard
& Trebbau
1984).
Ę
Nuchal scute
- a median scute that
lies over the nuchal bone anterior to the vertebral
scutes in many turtles (e.g.,
Pritchard &
Trebbau 1984).
Carr
(1952) called it the ōprecentralö but this has not
been accepted by others authors.
Zangerl
(1969) proposed the term "cervical scute" but
no previous authors and only a few subsequent ones
(e.g., Ernst
& Barbour 1989) have used this term. Nuchal
scute is usually small or absent in pleurodires (it
is wide in Hydromedusa).
Ę
Nuchaloid scute
¢ the small anterior
most marginal scute that lies over the lateral part
of the nuchal bone and anterolateral part of the
nuchal peripheral bone between the nuchal scute and
the marginal I. This term was proposed by
Chkhikvadze
(1973) but
has not been used
by others, who consider this scute to be the
marginal I. However, the nuchaloid scutes
distinguish from other marginal scutes by size and
anteromedial expansion in many pleurodires.
Ę
Vertebral scutes
are the (usually five)
large scutes along the midline of the carapace. This
term is nearly unanimous although
Carr
(1952) used ōcentralsö.
Ę
Pleural scutes
or pleurals are
the large scutes forming a longitudinal series on
each side of the vertebrals. This term was proposed
by Zangerl
(1969) and a number of authors have
subsequently adopted
Zangerl's
usage. Carr
(1952) used ōlateralö. These scutes are
termed ōcostalsö by
Pritchard &
Trebbau
(1984). This has also been accepted by other
authors. However, this term is more often used for
the underlying bones than for the scutes (see the
discussion under costals justifying use of this
term). I prefer to avoid usage ōcostalsö for scutes
because they have different origin and it is
necessary to distinguish between the underlying bone
of the axial skeleton and the superficial horny
scute. In comparison with the costal bones and
pleural scutes both the nuchal scute and nuchal bone
have dermal origin.
Ę
Marginals
are the numerous small scutes around the edge of the
shell, except those on the anterior (the nuchal and
nuchaloid scutes) and posterior (a supracaudal
scute) midline.
Ę
Supracaudals
are represented the two rearmost scutes of the
carapace (following
Pritchard &
Trebbau 1984). In some turtles
they are fused into a single scute.
Carr
(1952) proposed the alternative term ōpostcentralsö.
Some authors, including
Hay
(1908) and
Zangerl (1969), have simply considered these
scutes to be the posteriormost (normally the
twelfth) pair of marginals, and this is indeed how
they appear when they are paired. However, the
single large scute in this position (typical of most
tortoises, for example) has a different appearance
and according to
Pritchard &
Trebbau (1984) justifies a different
name.
2.2. Plastron
|
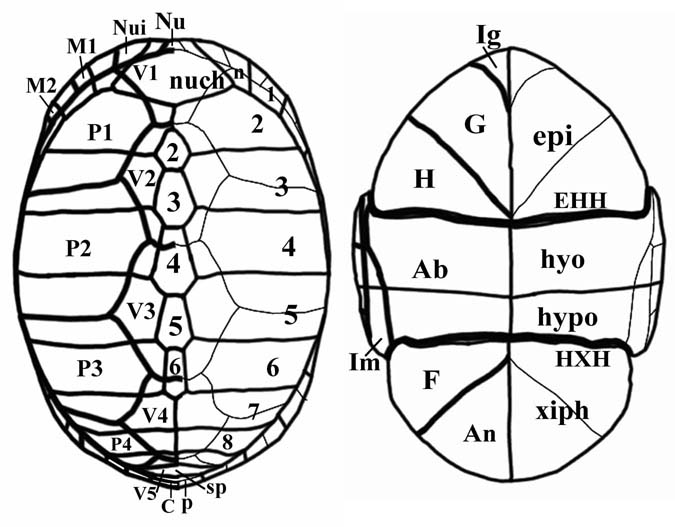
Kinosternon cruentatum (DumÉril & bibron 1851), Kinosternidae Abbreviations: Ab - an abdominal scute, An - an anal scute, cphf - a costoperipheral fontanelle, EHH - epihyoplastral hinge, ent - entoplastron; epi - epiplastron; F - a femoral scute, G - a gular scute, H - a humeral scute, hhf ¢ a hyohypoplastral fontanelle, hhxf - a hyohypoxiphiplastral fontanelle, hxf - hypoxiphiplastral fontanelle, hyo - hyoplastron; hyp ¢ hypoplastron, HXH - hypoxiphiplastral hinge, Ig - an intergular scute, Im - an inframarginal scute, M ¢ a marginal scute, n - a peripheral contacting with a nuchal, Nu - a nuchal scute, nuch - a nuchal, Nui - a nuchaloid scute, p - a pygal; SC ¢ supracaudal scute, sp - a suprapygal, xiph - xiphiplastron. |
Scutes of
the
plastron (not the
intergular, single gular, interanal, undivided anal,
etc.) are divided into pairs by a median
longitudinal seam. Anteriorly there is a pair of
gular scutes. In some families (notably
plesiochelyids, cheloniids, most kinosternids,
dermatemydids, pleurodires) an intergular scute
or pair of intergular scutes is also present. In
kinosternids
Hutchison &
Bramble
(1981) homologizate the intergular scute(s) with the
gulars, and the
posterior humerals with the abdominals
of other turtles
(see discussion in
Kordikova
2000, 2002). Paired humerals, pectorals,
abdominals, femorals, and anals
follow, respectively, and in the Cheloniidae an
interanal is sometimes present. Some tortoises
have two additional pairs of plastral scutes ¢
pregulars and preintergulars anterior to
the gulars and intergulars.
Ę Inframarginals - scutes between the main plastral scutes and the ventral edge of the marginal scutes. They are mainly present in Cheloniidae, Chelydridae, Dermatemydidae, Kinosternidae and Platysternidae. Inframarginals are often reduced to just the anteriormost and posteriormost members, known as the axillaries and inguinals, respectively. Some authors (Chkhikvadze 1973) use the term latiplastral.
3. Fontanelles
These structures are
not named by all authors even though fontanelles are
present at least in early stages of morphogeny in
almost all Testudinata. I distinguish
the following fontanelles.
3.1. Carapace
Ę Postnuchal fontanelle - fontanelle between the nuchal and anterior pair of costals.
It can be unpaired or
paired. Some authors (Meylan
1987,
Pritchard 1993) call it suprascapular one.
Ę
Neurocostal fontanelles
- fontanelles between costals and neurals.
Ę Intercostal fontanelles - fontanelles between neighboring thoracic ribs or costals.
Ę Costoperipheral fontanelles ¢ lateral fontanelles between costals and peripherals.
3.2. Plastron
Ę
Epihyohypoxiphiplastral fontanelle
¢ central fontanelle between epi-, hyo-, hypo- and
xiphiplastra. It is present in Dermochelys
coriacea.
Ę
Entohyoplastral fontanelle(s)
- unpaired or paired ventral fontanelles between
entoplastron and hyoplastra.
Ę
Entohyohypoplastral fontanelle(s)
- unpaired fontanelle between entoplastron, hyo- and
hypoplastra.
Ę
Hyohypoplastral,
or central fontanelle - fontanelle between
hyo- and hypoplastra. It is as a rule unpaired.
Ę
Hypoxiphiplastral fontanelle
or hyohypoxiphiplastral fontanelles - central
fontanelle between hypo- and xiphiplastra or between
hyo-, hypo- and xiphiplastra. It is unpaired. This
fontanelle is very often present in some
pelomedusids (e.g. Pelomedusa subrufa) or
adult cheloniids and trionychoids.
Ę
Xiphiplastral fontanelle
- unpaired fontanelle between xiphiplastra.
Ę
Plastroperipheral fontanelles
- paired lateral fontanelles between hyo-hypoplastra
and peripherals.
4. Hinges
Many turtles have
carapacial or plastral hinges (Borkin
1973,
Bramble 1974,
Bramble & Hutchison 1981,
Brumble et al.
1984,
Pritchard & Trebbau 1984,
Ernst & Barbour 1989,
Broadley 1993,
Pritchard 1993,
etc.).
These hinges allow the carapace or plastron to be
closed to protect head and limbs if they are large
and mobile enough. Hinges in chelonians can be
classified as:
4.1. Carapace
Ę
Costoperipheral hinge
- a transverse carapacial hinge that always lies
between the 5th and 6th
costals and peripherals or, sometimes, between the 6th
and 7th costals and peripherals. Most
species of the testudinid genus Kinixys only
has such a hinge.
Ę
Peripheral hinge
- a transverse
carapacial hinge between the 5th and 6th
peripherals in Kinixys natalensis.
Ę Carapacial and plastral pankinesis (Pritchard 1993) ¢ kinesis between all elements of the carapace and plastron ¢ is characteristic for chelonians having paedomorphic features. Among living species carapacial and plastral pankinesis is present in most trionychoids as well as in the cheloniid Eretmochelys imbricata. Plastral pankinesis is characteristic for cheloniids and some other chelonioids. In sea turtles the plastron has lines of flexibility at the bridge and the midline. Carettochelys insculpta has costoperipheral and plastroperipheral kinesis as well as the kinesis between the epi- and entoplastra, on the one hand, and hypoplastra, on the other hand. There is also a movable link between the hyo-, hypo- and xiphiplastra of the left and right sides. In most trionychoids the bony carapace is completely rigid, as is the hyohypoplastral suture line.
The carapacial and plastral pankinesis is associated with changes in shell depth and is the result of adaptive evolution within different groups.
4.2. Plastron
Ę Epihyoplastral hinge - a movable plastral hinge, which lies between the epiplastra and hyoplastra - between the humeral and pectoral scutes, either, crossing the posterior portion of the entoplastron or behind it. It is present in Pyxis (Testudinidae). In kinosternids this hinge lies across the humeral scute (following Bramle et al. 1984) or between the humeral and abdominal scutes because of reduction of the pectoral scute.
Ę
Hyomesoplastral hinge
¢ a transverse plastral hinge placed between hyo-
and mesoplastra. It is present in Pelusios (Pelomedusidae)
but it is rather rigid in P. broadleyi and
P. adansoni.
Ę
Hyohypoplastral hinge
¢ a transverse plastral hinge placed between the hyo-
and hypoplastra - between pectoral and abdominal
scutes. It occurs in many genera of emydids such as
Emys, Emydoidea, Terrapene, Cuora, Cyclemys,
Pyxidea, Notochelys and Rhinoclemmys.
Ę
Interhyo- and interhypoplastral hinge
¢ a longitudinal plastral hinge along the hyo- and
hypoplastral sutures (in Sternotherus spp.).
Ę
Hypoxiphiplastral hinge
¢ a transverse plastral hinge between hypo- and
xiphiplastra - between the abdominal and femoral
scutes. Hypoxiphiplastral hinge is well developed in
kinosternids. The plastron of kinosternids may
contain either an inconspicuous single
epihyoplastral hinge (e.g. Claudius, Staurotypus)
or two transverse movable hinges - epihyoplastral
and hypoxiphiplastral hinges (in most species of
Kinosternon). Most species of Testudo and
Gopherus (Testudinidae) have a weak
hypoxiphiplastral hinge.
Ę
Interxiphiplastral hinge
- a longitudinal
plastral hinge between the xiphiplastra (in
Kinosternon scorpioides).
Ę
Plastroperipheral hinge
- a longitudinal hinge
between the hyohypoplastra and peripherals of the
carapace (in Claudius angustatus and in
emydids - Emys orbicularis, Emydoidea spp.
and Terrapene spp., etc.)
Though this shell
nomenclature proposed here and earlier (Kordikova
2000) represents a relatively recent
innovation it seems to be logical and it has an
embryological and comparative morphological (homological)
basis for evolutionary applications.
5. Other terms
Ę
Seam
- a term for the line of connection between two
scutes.
Ę
Suture - a
term for the connection between two bones.
Ę Sulcus - a term for the impression that a seam leaves on the underlying bone, are of universal usage.
more information:
kordikova, E. G. (2000): Paedomorphosis in the shell of fossil and living turtles. - N. Jb. Geol. Palaont. Abh.,
Classification of lower tetrapods
Class Reptilia Linneus 1758
Subclass Parareptilia Olson 1947
Order
Procolophonomorpha
Suborder
Procolophonia
Superfamily
Procolophonoidea
Family Nyctiphruretidae Efremov 1938
Family Procolophonidae Seeley 1888
Suborder
Pareiasauroidea
Family Lanthanosuchidae Efremov 1946
Family Nycteroleteridae Romer 1956
Family Pareiasauridae Seeley 1888
Family Bradysauridae Huene 1948
Order Chelonia brongniart 1800
Suborder Proganochelydia ROMER 1966
Suborder Casichelydia GAFFNEY 1975
Subclass Eureptilia
Order Diadectida
Suborder
Diadectomorpha
Family Diadectidae
Order Captorhinida
Suborder
Captorhinomorpha
Family
Protorothyrididae
Family
Captorhinidae
Case 1911
Order Araeoscelida
Family
Petrolacosauridae
Order Choristodera
Family Champsosauridae
Infraclass
Lepidosauromorpha
Superorder
Lepidosauria
Order Sphenodontida
Family Sphenodontidae
Superorder
Sauropterygia
Order
Placodontia
Cope
1871
Suborder Placodontoidea Cope 1871
Family Placodontidae Cope 1871
Family Paraplacodontidae Peyer & Kuhn-Schnyder 1955
Suborder
Cyamodontoidea
Nopcsa
1923
Superfamily Cyamodontida Romer 1956
Family Henodontidae F.v. Huene 1948
Family
Cyamodontidae
Nopcsa
1923
Infraclass
Archosauromorpha
Superorder
Archosauria
Order Thecodontia
Suborder
Proterosuchia
Family Proterosuchidae
Suborder
Ornithosuchia
Family Euparkeriidae
Suborder
Aetosauria
Family
Stagonolepididae
|
Classification of fossil and living Chelonia |
Order Chelonia brongniart 1800 (Latreille 1800)
Suborder Proganochelydia Romer 1966
Family Proganochelyidae Baur 1888
Suborder Casichelydia GAFFNEY 1975
Infraorder Cryptodira Cope 1868
Family Pleurosternidae Cope 1868
Family Kallokibotionidae Nopcsa 1923
Family Solemydidae Lapparent de Broin & Murelaga 1996
Family Thalassemydidae Zittel 1889
Family Eurysternidae Dollo 1886
Family Plesiochelyidae Baur 1888
Superfamily Chelonioidea Oppel 1811
Family Osteopygidae Zangerl 1953
Family Protostegidae Cope 1872
Superfamily Dermochelyoidea Fitzinger 1843
Family Dermochelyidae FITZINGER 1843
Superfamily Trionychoidea Fitzinger 1826
Family Trionychidae Fitzinger 1826
Family Cyclanorbidae LYDEKKER 1889
Family Peltochelyidae Seeley 1880
Superfamily Carettochelyoidea Boulenger 1887
Family Carettochelydidae Boulenger 1887
Family Kinosternidae Agassiz 1857
Family Staurotypidae GRAY 1869
Superfamily Chelydroidea gray 1831
Family Platysternidae gray 1869
Family Dermatemydidae gray 1870
Superfamily Testudinoidea batsch 1788
Family Testudinidae batsch 1788
Superfamily Emydoidea Rafinesque 1815
Family Emydidae Rafinesque 1815
Infraorder Pleurodira COPE 1864
Family Proterochersidae Nopcsa 1928
Family Platychelyidae Brõm 1965
Family Dortokidae Lapparent de Broin & Murelaga 1996
Superfamily Pelomedusoides Cope 1868
Family Pelomedusidae Cope 1868
Superfamily Podocnemidoidea Cope 1868
Family Bothremydidae Baur 1891
Family Podocnemididae Cope 1868
Superfamily Chelidoidea Gray 1825
more information:
|
Classification of lower tetrapods Class Reptilia Linneus 1758 Subclass Parareptilia Olson 1947
Order Procolophonomorpha
Suborder
Procolophonia
Superfamily
Procolophonoidea Family Nyctiphruretidae Efremov 1938 Family Procolophonidae Seeley 1888
Suborder
Pareiasauroidea Family Lanthanosuchidae Efremov 1946 Family Nycteroleteridae Romer 1956 Family Pareiasauridae Seeley 1888 Family Bradysauridae Huene 1948 Order Chelonia brongniart 1800 Suborder Proganochelydia ROMER 1966 Suborder Casichelydia GAFFNEY 1975
Subclass Eureptilia
Order Diadectida
Suborder
Diadectomorpha
Family Diadectidae
Order Captorhinida
Suborder
Captorhinomorpha
Family Protorothyrididae
Family
Captorhinidae
Case 1911
Order Araeoscelida
Family Petrolacosauridae
Order Choristodera
Family Champsosauridae
Infraclass
Lepidosauromorpha
Superorder Lepidosauria
Order Sphenodontida
Family Sphenodontidae
Superorder Sauropterygia
Order
Placodontia
Cope
1871 Suborder Placodontoidea Cope 1871 Family Placodontidae Cope 1871 Family Paraplacodontidae Peyer & Kuhn-Schnyder 1955
Suborder
Cyamodontoidea
Nopcsa
1923 Superfamily Cyamodontida Romer 1956 Family Henodontidae F.v. Huene 1948
Family
Cyamodontidae
Nopcsa
1923
Infraclass
Archosauromorpha
Superorder Archosauria
Order Thecodontia
Suborder
Proterosuchia
Family Proterosuchidae
Suborder
Ornithosuchia
Family Euparkeriidae
Suborder
Aetosauria
Family Stagonolepididae
Order Chelonia brongniart 1800 (Latreille 1800) Suborder Proganochelydia Romer 1966 Family Proganochelyidae Baur 1888 Suborder Casichelydia GAFFNEY 1975 Infraorder Cryptodira Cope 1868 Family Pleurosternidae Cope 1868 Family Kallokibotionidae Nopcsa 1923 Family Solemydidae Lapparent de Broin & Murelaga 1996 Family Thalassemydidae Zittel 1889 Family Eurysternidae Dollo 1886 Family Plesiochelyidae Baur 1888 Superfamily Chelonioidea Oppel 1811 Family Osteopygidae Zangerl 1953 Family Protostegidae Cope 1872 Superfamily Dermochelyoidea Fitzinger 1843 Family Dermochelyidae FITZINGER 1843 Superfamily Trionychoidea Fitzinger 1826 Family Trionychidae Fitzinger 1826 Family Cyclanorbidae LYDEKKER 1889 Family Peltochelyidae Seeley 1880 Superfamily Carettochelyoidea Boulenger 1887 Family Carettochelydidae Boulenger 1887 Family Kinosternidae Agassiz 1857 Family Staurotypidae GRAY 1869 Superfamily Chelydroidea gray 1831 Family Platysternidae gray 1869 Family Dermatemydidae gray 1870 Superfamily Testudinoidea batsch 1788 Family Testudinidae batsch 1788 Superfamily Emydoidea Rafinesque 1815 Family Emydidae Rafinesque 1815 Infraorder Pleurodira COPE 1864 Family Proterochersidae Nopcsa 1928 Family Platychelyidae Brõm 1965 Family Dortokidae Lapparent de Broin & Murelaga 1996 Superfamily Pelomedusoides Cope 1868 Family Pelomedusidae Cope 1868 Superfamily Podocnemidoidea Cope 1868 Family Bothremydidae Baur 1891 Family Podocnemididae Cope 1868 Superfamily Chelidoidea Gray 1825
|